
ギムザ染色法
放射線被曝線量の生物学的評価:ギムザ染色法による安定型染色体異常の検出法
阿波 章夫
目次
- はじめに
- 染色体標本の作成
- 分裂期細胞(メタフェース)の選び方
- 染色体表記法
- 各群染色体の特徴(核型分析)
- 正常変異染色体の種類
- 安定型構造異常-総論
- 安定型構造異常-各論1.欠失
- 安定型構造異常-各論2.逆位
- 安定型構造異常-各論3.相互転座
- 誤った判定の例
- 終わりに
第1章.はじめに
血液リンパ球を培養して染色体を観察し、その異常頻度を測定すると各人が受けた放射線量を推定できます。染色体異常の中には、不安定型異常と、安定型異常の2種類があります。
不安定型異常とは、2動原体染色体や環状染色体のことで、観察は容易ですが、細胞分裂に伴って異常を持つ細胞が失われていくという性質があります。従って被曝後何年もの時間を経てしまうと頻度が低くなってしまうという問題があります。他方の安定型異常は、相互転座や逆位染色体のことで、細胞分裂による影響を受けません。従って被曝後長い年数を経ても頻度が低下することはないようです。しかしこの安定型異常は観察が容易でなく、検出感度が低いという欠点がありました。
原爆被爆者の染色体異常頻度に関する調査は1960年代後半に開始されましたが、調査開始時点ですでに不安定型異常はほとんど消滅していました。従って安定型異常を調査するしか方法はありませんでした。当時は普通のギムザ染色しか方法はありませんでしたから、試行錯誤して異常検出のマニュアルを作りました。そして約30年間にわたって調査を行ってきました。
最近になって従来のギムザ染色に代わって染色体着色法(FISH)が利用できるようになりました。この方法は、特定の染色体に色をつけることにより、異なる色の染色体間の転座を客観的に検出できます。200名以上の原爆被爆者について、従来のギムザ法とFISH法の結果を比較してみたところ、従来のギムザ法では転座の約70%を検出していたことが明らかになりました。 (Nakano M et al. Int J Radiat Biol 77:971-7, 2001)
この手引書を公表することにした理由は、最近ではFISH技術に余りに多くの眼が向いてしまって、あたかもそれ以外の方法では安定型染色体異常の検出はできないという風潮が強くなったので、異なる考えを示したかった次第です。確かにFISH法には多くの利点がありますが、最大の欠点はコストが高いことと高価な蛍光顕微鏡を必要とする点です。そこで最もコストの低いギムザ染色という方法でも、研究者の姿勢次第ではかなりのレベルの調査が可能であることを示したいと思います。
第2章.染色体標本の作成
ここでは染色体分析のための血液培養法とか、顕微鏡標本作成法などの技術面については触れません。以下の専門書を参考にして下さい。
外村 晶編; 「染色体異常」朝倉書店、1978年
古庄敏行編; 「臨床染色体診断法」金原出版、1996年
Verma RS and Babu A; Human chromosomes. Manual of basic techniques. New York, Pergamonn Press, 1989.
第3章.分裂期細胞(メタフェース)の選び方
ギムザ染色の場合は、緑(または黄色)フィルターを用います。最もコントラストがよく、写真撮影にも適しています。
まず低倍率レンズを用いてメタフェースを選びます。大切なことは、「低倍率で選んだメタフェースのすべてが高倍率における分析に使える訳ではない」ことです。ここでいう低倍率とは、10(接眼レンズ)×10(対物レンズ)=100倍、あるいは10×20=200倍であり、高倍率とは 10(または15)×100(油浸レンズ、もしくはドライ・レンズ)=1000(あるいは1500)倍を指します。低倍率で選んだメタフェースに自信がない時には、中程度の倍率(400倍:10×40)で確かめることを薦めます。
良いメタフェースの条件は:
- 46本の染色体が同一平面上に均一に、かつ同心円状(やや楕円状でも良い)に広がっていること。
- 染色体同志の重なりがないこと。たとえ、あったとしても2~3本程度の重なりに限られていること。
- 動原体が、全ての染色体においてはっきり確認できること。染色体が重なっていても動原体は重なっていないことが必要。
- 染色体の染まりが一定であること。つまり、どの染色体も同じ濃さで染められていること。一つのメタフェースの中で、左右または上下の染色体の染まり方に“むら”がある場合は、染色体の長さが不揃いを生じているので、このようなメタフェースを選ぶことは避ける。これは、空気乾燥法の時に生じた“ひずみ”であり、相同染色体間でも長さが違うことがある。一般的には、染色が薄ければ染色体は細長く、染色が濃ければ染色体は太く短い。
- 染色体数が46であることが望ましいが、低倍率での数の算定は難しい。染色体数の“ぶれ”は46(+-)1程度。
- 染色体を構成している2本の染色分体が明瞭に観察できること。
- コルヒチンの影響が強すぎて染色体が極度に短縮したもの、逆に染色体のらせん(螺旋)構造が強調されすぎているものは避ける。どちらも動原体の位置が不明瞭となる。
これら7条件のうち、低倍率で判定できるのは 項目(1)くらいで、他の条件は中、高倍率の観察によってはじめて判定が可能です。メタフェースの中で分析に適したもの、適していないものについて、図1から図3に示す顕微鏡写真によるメタフェースについて検証してみます。これらはいずれも 高倍率(10×100=1000倍)で撮影したものです。

図1.
図1.A~D は観察に適したと判断されるメタフェースです。その中でも、図1-A は重なりもなく、円状に程よく広がった良いメタフェースです。図1-B や 図1-D はAと比べて染色体が全般的にやや短い傾向にあります。これは分裂中期でも終わりに近い時期にあることと、染色体を短縮させる効果をもつコルヒチンの作用がやや効きすぎたためと思われます。図1-C も大変良いのですが、よく見ると、点線の矢印で示してあるように、2本の染色体が並列して、1本の異常染色体(2動原体染色体)のように見える例です。
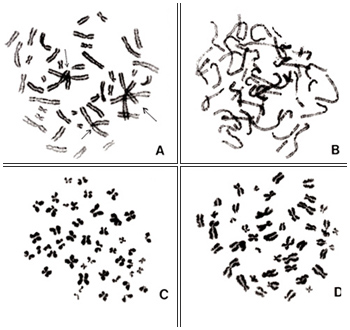
図2.
染色体分析に適さないメタフェースを 図2~3 に示します。理由は次の通りです。
図2-A は重なりが多いこと(矢印)、しかも、動原体同志の重なりにより動原体の正しい位置を判定できないため分析は不可能です。
図2-B は分裂中期というよりは分裂前期の染色体であり、個々の染色体を識別できないのが理由です。
図2-C~2-D は、いずれもコルヒチンが効きすぎているため、染色体はすべて短縮しすぎ、動原体の位置も不明瞭です。図2-D の中には折れ曲がった染色体もあります。こういう染色体は長さを正しく読み取ることができません。
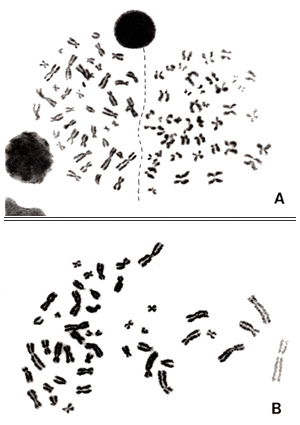
図3.
図3-A は2つのメタフェースが隣り合っているケースです。
この例では、両者の境界線はまだよくわかりますし、分裂の進行状況も違うので、両者を区別するのは容易です(点線)。しかし、2つのメタフェースが横並びに見える場合に注意しなければならないのは、一度分裂して新たに作られた2つの細胞(娘細胞)が、並び合ったまま同時に2回目の分裂に入る場合があることです。その場合、両者の染色体は、凝縮状態はウリふたつのままで部分的にまざり合うので、誤った判定をする恐れがあります。
図3-B は1つのメタフェースで拡がり具合が左右で異なることです。左側は濃く染まっていますが、右にゆくにつれて染まりは薄く、染色体も長くなります。伸びた側の染色体を異常と判定しかねないので、分析に採用するのは見合わせるべきでしょう。
低倍率で選ばれたメタフェースについては、高倍率レンズ(油浸レンズ)に切り替えて観察・分析を行います。その概要は以下のとおりです。
- 最初に、染色体数を数えます。慣れないうちはカウンターを使っても構いません。 しかし、じきに慣れて頭の中でカウントできるようになります。合格基準は、45、46、47本の場合だけです。
- 次に個々の染色体の形や長さについて分析を行います(核型分析といいます)。通常の方法は、2000~3000倍に拡大した顕微鏡写真から個々の染色体を切り出し、ルールにしたがって相同染色体を対に並べて、異常の有無を確かめ、もし異常があればどの染色体が異常なのかを判定します。
- しかし、これでは現実には異常の判定に時間が掛かりすぎるので、一歩進めて、顕微鏡下で直接メタフェースを観察しながら異常の有無を判定するための訓練が必要です。これができるようになれば、正常なことが確実な場合、顕微鏡写真を撮って染色体を切り出す必要はなくなります。ただし、明らかに異常が存在する場合はもちろんのこと、少しでも異常が疑われる場合は、必ず写真撮影することが大切です。
第4章.染色体表記法
46本の染色体にはヒトに特有の規則性が認められます。この規則性をルール化した「ヒト染色体に関する国際命名規約(ISCN)」があり、染色体分析はこのルールに従って行います 。(註1)
この中には、染色体とその部分、および染色体の異常などを表記できるさまざまな略称やシンボルのリストがあります。重要と思われる項目を 表1. に示します。
表1. 安定型染色体を表記するための主要なシンボルと略記法について |
|||
|
| cen | centromere | 動原体(セントロメア) |
| cx | complex interchange | 複合交換型異常 |
| del | deletion | 欠失 |
| h | heterochromatin | 異質染色質(ヘテロクロマチン)=二次狭窄 |
| ins | insertion | 挿入 |
| inv | inversion | 逆位 |
| mar | marker chromosome | どの染色体群にも属さない異常染色体 |
| – | loss | 消失、または喪失 |
| p | short arm of chromosome | (染色体の)短腕 |
| pcd | premature centromere division | 動原体の早期分裂 |
| + | gain | 増加 |
| q | long arm of chromosome | (染色体の)長腕 |
| rcp | reciprocal | 相互型(転座など) |
| s | satellite | 付随体 |
| ; | separate altered chromosomes and breakpoints |
二個(以上)の異常染色体を切り離して記載 |
| t | translocation | 転座 |
メタフェースの観察を始める前に染色体を構成しているさまざまな部位の名称の中で、特に重要と思われる項目について以下に記述します。すべての染色体には必ず動原体(cen)があり、cenを境にして長い部分を長腕(q)、短い部分を短腕(p)と呼びます。核型分析では、短腕を上、長腕を下に位置します。
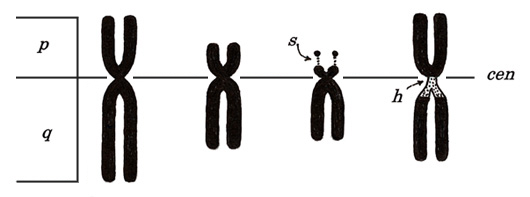
図4.
右から2番目の図に示すように、13番、14番、15番、21番、22番染色体の短腕の先端にはカタツムリの眼のような付随体(satellite: s)が存在します。ここにはリボソームRNAの遺伝子が存在しています。“h”の記号で示すものは二次狭窄とよばれるものです。一次狭窄はcenを意味し、これはすべての染色体に存在します。“h”は特定の染色体に出現し、1番、9番、16番染色体においてはcen近くの“くびれ”状の構造として観察され、薄くて弱い染色性を示します(ヘテロクロマチン)。他方Y染色体においては、“h”はcenではなく長腕の末端部に観察されます。
個々の染色体の特徴を理解するためには、表2. を見てください。長腕と短腕の長さを計測し、全長に対する短腕の比をセントロメア比(CI = centromere index)と呼びます。比の値が 50に近づくにつれて、cenは中央部にありますが、値が小さくなるにつれて、cenは末端部に位置するわけです。
表2 染色体の長さ (%による相対値: *1) と セントロメア比
(*2) |
|||
|
1
|
9.11 ( 4.43:4.68 ) | 48.6 |
2
|
8.61 ( 3.35:5.26 ) | 38.9 |
3
|
6.97 ( 3.30:3.67 ) | 47.3 |
4
|
6.49 ( 1.80:4.69 ) | 27.8 |
5
|
6.21 ( 1.66:4.55 ) | 26.8 |
6
|
6.07 ( 2.30:3.77 ) | 37.9 |
7
|
5.43 ( 2.01:3.42 ) | 37.0 |
X
|
5.16 ( 1.94:3.22 ) | 37.5 |
8
|
4.94 ( 1.62:3.32 ) | 32.8 |
9
|
4.78 ( 1.56:3.22 ) | 32.7 |
10
|
4.80 ( 1.55:3.25 ) | 32.3 |
11
|
4.82 ( 1.95:2.87 ) | 40.5 |
12
|
4.50 ( 1.23:3.27 ) | 27.4 |
13
|
3.87 ( 0.64:3.23 ) | 16.6 |
14
|
3.74 ( 0.69:3.05 ) | 18.4 |
15
|
3.30 ( 0.58:2.72 ) | 17.6 |
16
|
3.14 ( 1.33:1.81 ) | 42.5 |
17
|
2.97 ( 0.94:2.03 ) | 31.9 |
18
|
2.78 ( 0.74:2.04 ) | 26.6 |
19
|
2.46 ( 1.10:1.36 ) | 44.9 |
20
|
2.25 ( 1.03:1.22 ) | 45.6 |
21
|
1.70 ( 0.49:1.21 ) | 28.6 |
22
|
1.80 ( 0.51:1.29 ) | 28.2 |
Y
|
2.21 ( 0.51:1.70 ) | 23.1 |
| *1 常染色体(22対44本)の長さの総和を分母としてパーセンテージで表す。 *2 セントロメア比 (centromere index) = [ 短腕 ÷ (短腕 + 長腕) ] × 100 |
筆者註
- 註1:
- 染色体研究の専門家が1960年に米国デンヴァーに集まり、人類染色体国際命名規約委員会を発足させ、規則性に基づく染色体の分類法についてルール作りが行われました。これはデンヴァー会議と呼ばれ、人類染色体研究に関する統一された公式の国際的規約が作られたという画期的なイベントでした。この後、1963年にロンドンで新知見を取り入れたルールの改訂が行われました(ロンドン会議)。さらに、国際人類遺伝学会議が開かれる度に改訂や追加作業が行われることになり、1966年にはシカゴで、また1971年にはパリで、それぞれ会議が開かれました。
1976年にメキシコで国際人類遺伝学会議が開かれた時、世界の各地域を代表する命名規約国際常任委員を選挙によって選出することが決まりました。それまでは、規約がデンヴァー会議、ロンドン会議、シカゴ会議、パリ会議などのように、会議が開かれた地名を付けて呼ばれていましたが、1978年以後、規約は[人類染色体国際命名規約(An International System for Human Cytogenetic Nomenclature)]の頭文字のISCNとし、それに改訂の年度をつけることになりました [ISCN(1978)、ISCN(1981)、ISCN(1985)、ISCN(1991)、ISCN(1995)]。さらに、それぞれには、新しい技法の進展に基づく新しいルールの内容を表わす副題が付記されます。ISCN(1981)には高精度分染法、また、ISCN(1991)には癌の染色体のためのガイドラインが記載されました。
第5章.各群染色体の特徴:核型分析

図5.
核型分析(karyotype)の原則は、22対の常染色体を長さの順に、そして性染色体を最後に並べます。長さとcenの位置を基準にして染色体を並べれば、特徴的な7つの染色体群(A~G)に区別できます。それぞれの群の特徴について以下に述べます。
図5. にヒト染色体パターンの模式図(ideogramまたはidiogram)を、また図6. に写真分析による男性と女性のメタフェースの核型分析(karyotype)の結果を示しました。以下に述べる特徴や定義の理解に役立てて下さい。(註2)
先に示した 表2. はISCN(1985)からの引用ですが、もとの表にあった観察の標準誤差の値は省略しました。誤差の範囲は 約5~10%程度であることを念頭に置いて利用してください。なお、C群染色体の識別がもっとも困難なので、説明は最後にすることにします。
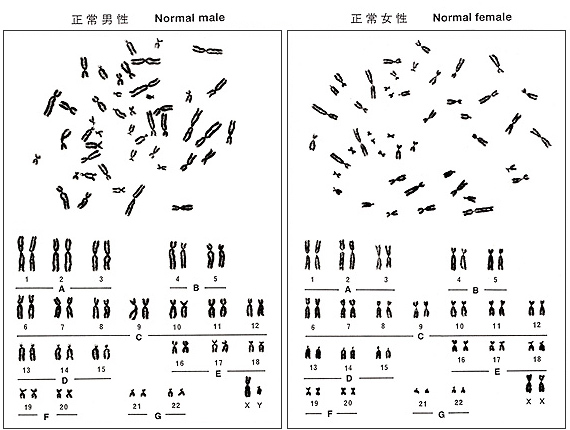
図6.
A群 (染色体1~3)
いちばん大型の3対の染色体でcenの位置はいずれも中央部にあります。1番と2番の長さはほぼ同じですが、cenの位置が違います。1番はほぼ中央にありますが、2番は短腕が長腕よりも短く、4:6の割合でcenの位置が偏っています。つまり、CIは 約40%ということです。1番には第二次狭窄“h”があるので、2番とは容易に区別ができます。3番は1番と同じくcenの位置が中央部にあり、形も1番に実に似ています(1番=48.6%: 3番=47.3%)が、3番のほうが20%程短いため、慣れるにしたがって識別は容易となります。
問題は 1番の“h”です。相同染色体の両方にhがある人もあれば、どちらか一方だけに見られる人もあります。人によってはhが全く見られない場合もあります。1番のhは常にcenに続く長腕上部にあります。その形を一言で表現すると、長い首に続いて、“なで肩”のように見えます。まるで、丹頂鶴の首から羽のあたりを見ているようです。 図6 左(男性)の右側1番染色体の形を参照してください。ヘテロクロマチンの量が多いと“h”の範囲は長くなります。これは構造的異常とは考えられてはいません。正常変異(normal variant)、あるいは異形染色体(heteromorphic chromosome)と呼ばれる現象です。親のいずれかがこの変異を保有している場合は、メンデルの法則によりその子供の半分に発現します。
B群 (染色体4~5)
大型の2対の染色体でcenは末端部寄りにあります。B群の特徴は、A群に次いで長く(1番の全長の 約75%程度)、CIは 27~28です。ということは、全長の4分の1が短腕という訳です。B群の長腕の長さは1番の長腕の長さにほぼ匹敵します。4番と5番は長さ、形ともに非常によく似ているため、通常染色法の標本では両者の識別はできません。したがって、4本を一括してB群として扱います。
C群 (後述)
D群 (染色体13~15)
中型の染色体3対からなり、全長は3番染色体の短腕に相当します。末端部にcenが存在し(CI=17~18)、全部の染色体の短腕に“s”が存在します。G群染色体と共通した特徴的な形態のため、他の染色体との区別は容易です。 “s”は標本のでき具合によっては全く見られないこともありますが、ほとんどの場合は観察できます。メタフェースにおいては、D、G群の染色体の“s”の部分が集合して 付随体連合(satellite association)を形成することがあります。これはリボソームRNAの合成と関係があります。
“s”の大きさや形態に関しては“h”と同様に正常変異と考えられる個人差が存在し、メンデルの法則に従って遺伝します。稀には短腕の大部分が欠落している場合もありますが、遺伝性疾患とは関係がありません。
E群 (染色体16~18)
D群よりやや小型の3対の染色体です。cenは、16番染色体はほぼ中央部(CI=42)に位置しますが、17番(CI=32)、18番(CI=27)は端部寄りに位置し、短腕が明らかに短い傾向にあります。17番と18番は長さも形も非常によく似ていますが、短腕の長さが 17番よりも18番の方が少し短いので、経験を積むと顕微鏡下で識別ができます。16番には1番と同じ場所に顕著な“h”が観察されることがあり、形と長さに個人差が見られます。
F群 (染色体19~20)
2対の短い染色体。cenがほぼ中央部にあり、19番と20番の識別はできません。一括してF群染色体として扱います。
G群 (染色体21~22, Y)
最小の2対の常染色体とY染色体から成るので男性では 5本、女性では 4本観察されます。形態的特徴はD群と同じで、cenは末端部にあり、すべての短腕に“s”があり異形染色体としての個人差がみられます。 しかしYには“s”は存在しません。21番の方が22番よりも小さいことが後で判明しましたが、最初に取り決めた核型を採用しています。
Y染色体は 21番や22番よりもやや長めです。長腕の下半分位はヘテロクロマチンでできており、正常変異としての長さの変化が多いのがYの特徴です。通常染色法では、“h”の部分の染色は薄く、その部位の染色分体は離れずにくっついているように見えます。さらに、“s”が存在しないことなどから、他のG群とは容易に識別することができます。
C群 (染色体6~12, X)
中型の染色体で7対の常染色体とX染色体からなるため、男性では 計15本、女性では 計16本が含まれます。cenの位置は中央寄りかやや末端部寄りです。X染色体はこのグループの中の大きい方に属します。このグループの染色体は、いずれも長さや形が互いに似ているので、それぞれを識別することはかなり困難です。そういう難しさは存在しますが、通常染色法でも指摘できるC群の個々の染色体の客観的な特徴を提供したいと思います。
長さの順に染色体を並べて見ると、C群の特徴の一つはcenの位置にあります。つまり、CI値がわずかではあるが互いに異なっています。理解を助けるために少々誇張して表現してみると、6番: CI=大、7番+X: CI=大、8番: CI=小、9番: CI=やや小、10番: CI=小、11番: CI=大、12番: CI=小、という傾向になります。6番から12番までは、大/大/小/やや小(hあり)/小/大/小、の順になります。並べてみると、cenの位置がデコボコになります。
6番のペアが一番大きく、並べてみると7番よりも大きいことが分かります。しかしメタフェースのままで、このわずかな長さの違いを指摘するまでにはかなりの経験を必要とします。6番短腕の中間部にはしばしば染まりの薄い部位が認められます。
7番とXを区別するのはほぼ不可能です。従って、C群では最大の6番に続く大きめの3本(男性)または4本(女性)のCI=大に当てはまる染色体を7番+Xと定義して並べます。
X染色体に関しては、50歳を過ぎた女性においては細胞によっては2本のXのうちのどちらか一方にcenの“くびれ”構造が認められない場合があります。即ち2本の染色分体が、あたかも染色体断片のように並んで見えます。これは不活性化されているX染色体に由来するもので、老齢の女性に多く観察され、年齢の増加と共にその頻度も上昇する傾向があります。
8番、10番、12番のcenの位置はやや末端寄りという以外には、あまり特徴はありません。特に10番と12番は非常によく似ていますから、識別は困難です。便宜的に最小の対を12番の位置に並べます。
これらの中で、9番のペアの特徴は比較的容易に認識できます。それはこの染色体には1番、16番と同様に二次狭窄“h”が存在し、個人差を示す場合がしばしばあるからです。前にも述べたように、“h”の形態は、鶴の頭をcenとすれば、「首」とそれに続く「羽部」に至る“なで肩”のような構造という表現がピッタリです。こうした異形染色体はかなりの高頻度で特定の家系に観察できます。
9番のペアに見られるもう1つの正常変異は、約100人に1人の割合で発見される逆位(短腕と長腕が入れ替わっている構造異常)です。これはメンデルの法則に従って遺伝するので、両親のいずれかが保因者であれば子供の半分には9番ペアの一方が逆位になっています。しかし逆位があったからといって病気との関連は認められないので正常な変異体のひとつと考えられています。
以上がそれぞれの群における染色体の特徴です。この識別の最善の方法は、AからG群に属する染色体数をそれぞれ正確に把握することなのです。女性なら: A=6、B=4、C=16、D=6、E=6、F=4、G=4であり、男性なら: A=6、B=4、C=15、D=6、E=6、F=6、G=5であることを確認できるように訓練することです。逆に、性別不明の標本を観察した結果、前述後半の染色体数を得たなら、あなたは男性の細胞を観察していたのです。
要約しますとここで大事な点は: 染色体の正常・異常を確認し、決定するには、A群からG群の染色体の数を正確に数え、そして、それぞれの群に余計な染色体がなければ正常なのです。だから、もし、どこかの染色体群の数が多く、別の群で少ないという場合には、異常染色体が必ずどこかに存在すると疑うわけです。
何もこの順番通りに染色体を数える必要はありません。私の場合は、D群、G群、F群、E群(16番と17-18番)、A群、B群の順の観察を行い、最後に丹念にC群について数を数えながら分析を行います。
個々のグループについて数える時に、メタフェースの染色体をスケッチする習慣をつけることを是非薦めます。慣れてくるとさほどの時間はかかりませんし、後日、観察した細胞の証拠にもなります。
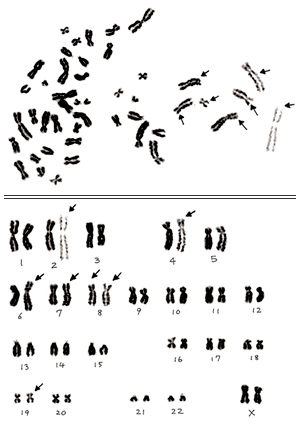
図7.
第3章の 図3-B において、部分的にゆがみがあって不適当と思われる細胞を示しました。このメタフェースとそれから作成したカリオタイプを 図7. に示します。
この図からも明らかな通り、メタフェースの右側の染色体はゆがみのために異常に伸長しています。右端の数本の染色体はらせん(螺旋)構造が観察できるほど極端に伸展しています。特に矢印をつけた染色体は相同のパートナーと比べて短腕・長腕共に伸びすぎています。カリオタイプから推定して、この細胞は正常であろうと考えられますが、これを採用するのは危険であり、カリオタイプをする以前の段階で棄却すべきです。
筆者註
- 註2:
-
際命名規約では 核型分析(karyotype) と 模式図(ideogramまたはidiogram) をはっきり区別しています。両者は染色体パターンを表現するものとして無差別に用いられていますが、厳密には少し違います。
図6 に示すように、たとえ1個のメタフェースを対象としたものであっても、描写、写真、もしくはイメージ・アナライザーによる染色体の配列・分析の結果から得られたヒト(もしくは特定の生物種)の染色体パターンを示すものを核型分析と呼びます。一方、模式図とは多数のメタフェースの分析と測定の結果を模式化し、ヒト(もしくは特定の生物種)の染色体パターンを図式化したものを指します。
第6章.正常変異染色体の種類
第5章で説明した個人差について例を示します。もしもこういう人の染色体異常を観察することになっても、全ての細胞に同じ変化が認められるので放射線によって生じた異常とは簡単に区別できます。
染色体の変異(正常変異、あるいは異形染色体)を生まれながらに持つ人がヒト集団中に 2%~5%の割合で存在します。ほとんどが“h”と“s”の変異に関するもので例を 図8. と 図9. に示します。
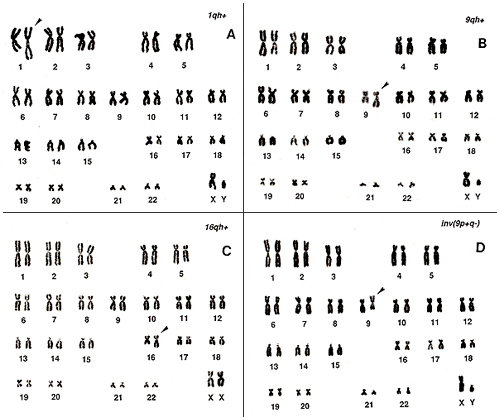
図8.
図8-A では1番、Bでは9番、Cでは16番の“h”が細長く伸びた状態になっており、1個の細胞だけを観察すると染色体の形態的異常と判定してしまう恐れがあります。特に、図8-C の16番では長腕が“鶴の首”状を示さなければ、C群の 10番や 12番と全く区別がつきません。
図8-D は 9番染色体が逆位を示す例です。この異常は親から受け継がれた遺伝的な形質です。この異常自体は自然集団中に約1%という比較的高い頻度で存在し、病気との関連はありません。

図9.
図9. はD群とG群の短腕、もしくは“s”に関連する正常変異体です。これには短腕もしくは“s”部位が大きくなっている場合(図9-A はD群、図9-B はG群)と、短腕が完全に欠失した場合(図9-C はD群、図9-D はG群)とがあります。いずれも識別は容易です。
第7章.安定型構造異常-総論
基本的な姿勢は、染色体の長さとCI値、あるいはhとs部位の長さなどを用いて、正常と異常の間の微妙な違いをできるだけ客観的に判定することです。
同じ長さの異常でもそれが長い染色体に生じた場合には検出しにくく、反対に短い染色体に生じた場合には検出しやすいという事実があります。8マイクロメートルの染色体が 10マイクロメートルに変化しても、その違い(25%増)を簡単に決めることはできませんが、同じ2マイクロメートルの増加でも、それが 2マイクロメートルの染色体に生じたなら(つまり 4マイクロメートルに変化した場合)は、その違いは容易に指摘できます。
染色性には変化はないけれど、相同染色体の一方が 10%長い時は: (i)一方が短腕・長腕共に 10%ずつ伸びている場合と、(ii)短腕(または長腕)の長さは同じだが、長腕(短腕)が全体の 10%に匹敵する分だけ長い場合、の2通りの可能性があります。(i)はCIに変化はなく、(ii)はCIが変化していることになります。この場合の常識的な解答は (i)は正常の範囲内での変化であり、(ii)は恐らくは異常染色体であろうと予想されます。長さの違いだけではなく、CIの違いの有無を確認することが大切です。
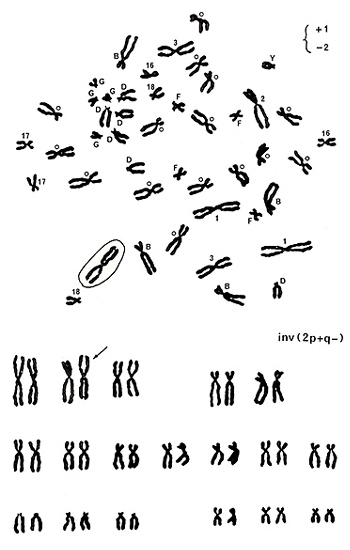
図10.
その他にメタフェースの時期、つまり、中期の前期か後期かによっても、また、1本の染色体の中でも、部分的な幅の違い、あるいは染色性が薄いか濃いか、によっても影響されます。同一メタフェース中の相同染色体の間でも、染色性が濃いものと薄いものがあります。濃く染まる染色体は太く短くなる傾向がありますし、染色性が薄い染色体は細長くなります。つまり、染色体量=染色の濃さ(太さ)×長さ=一定、ということです。この関係はほとんどの場合に当てはまります。
では安定型異常検出の具体的な方法に入りましょう。 図10. 上には染色体のcenの付近にいろいろな記号が書き添えてあります。数字の1、2、3、16、17、18は染色体番号であり、B、D、F、Gは染色体群を表わします。C群には○印がつけられており、Y染色体はそのままYを用いています。慣れてくると、Yの判定がもっとも容易だからです。
Y、G群、D群、16、17、18の数はそれぞれ正常です。B群は4本、C群は15本あり、いずれも変な染色体は見当たらないので、これらも正常のようです。1、2、2を見ると、3番は正常ですが、1番が3本と2番が1本であることが確かめられました。図10 の右上に+1、-2と記されています。1番に見えた3本の中の1本を丸で囲いました。なぜならば、この染色体には1番に特有の“h”がcenの付近にないからです。
このメタフェースにある1本の染色体が異常のようで、他の45本は正常と判断されます。実際にカリオタイプして見ると(図10下)、2番染色体の逆位(inv[2p+q-])であることが分かります。このように、メタフェースの中の1本だけが異常な時は、欠失か逆位が疑われます。
以下それぞれの安定型異常について例を説明していくことにしましょう。
第8章.安定型構造異常-各論1.欠失
欠失delは1本の染色体の長腕か短腕のいずれかが短くなっている異常です。僅か1本の異常ですから、逆位との区別が難しい場合があります。また逆位と同様に長い染色体に多く出現する異常でもあります。
4つの異なるメタフェースとカリオタイプによる具体例を 図11、12 に示します。これらの図に示す異常もA群やB群で占められています。一方、小さな染色体では、ほんのわずかの欠失でも識別できますから、異常の発見率が逆に偏る可能性があります。
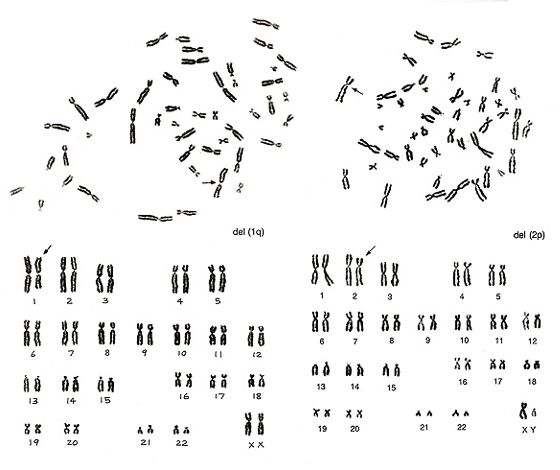
図11.
図11左は、B群からG群までに異常はありませんが、A群に異常があります。1番が1本なのに対して、3番が3本あります。この結果から、1番染色体の長腕の欠失del(1q)が予測されます。図11右は2番染色体が1本足りず、B群染色体によく似た異常染色体があります。よく見ると、短腕も長腕もB群よりもわずかに長いこと、および長腕の長さが2番とほぼ同じであることから、2番染色体短腕の欠失del(2p)と判定されます。
図12. 左右は異常の判定が難しい例で、いずれもB群染色体が関与しています。
左の図は、矢印で示すB群の1本にやや異常が疑われます。残りの3本よりも短腕が心持ち短い程度です。もしも、長腕が他の3本よりもやや長ければ、おそらくは逆位が疑われるところです。長腕は他のB群染色体と比べて長いわけでもないので、ここは短腕部位の欠失と判定しました。この判定の唯一の基準はCIが 20%か、それ以下という点にあります。
右図もB群染色体の異常です。4本のうちの1本がC群染色体の中でも小さい方に属するものと区別できない程類似しています。つまり、カリオタイプはC群が1本多く、B群が1本少ないことになります。記載にある通り、del(Bq)、つまりB群染色体長腕の欠失と判定されます。
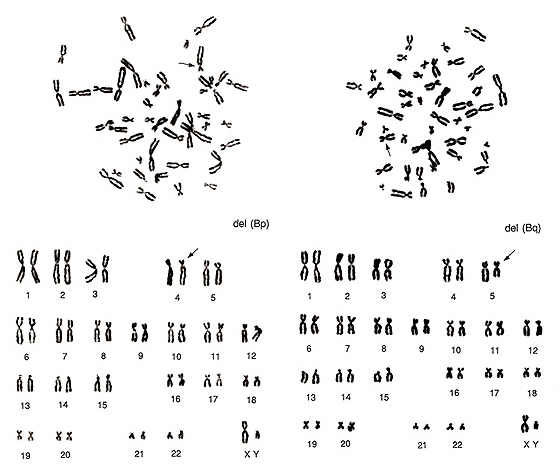
図12.
欠失型異常を識別するポイントは、1つの染色体群の中で1本だけが異常であることで、ある染色体群の染色体が1本不足していて同時に他のどの染色体群にも属さない形や長さの異常を生じる場合と、1ランク下の染色体群に存在する過剰な染色体として発見される(つまりB群染色体の欠失は1ランク下のC群染色体1本の増加となる)場合とがあります。
第9章.安定型構造異常-各論2.逆位
逆位の特徴はどの染色体群にも属しない異様な形の染色体が1本だけ出現し、その結果としてある染色体群の染色体数が1本不足することです。このような特異な形態異常の染色体をマーカー染色体と呼びますが、これは欠失や転座にも見られます。逆位の特徴は、ある染色体群に属する染色体が逆位の場合、逆位染色体の長さはその群に特有の長さの範囲を越えませんがCIに変化を生じます。長さに関する限りは各群の染色体数に変化はありません。
cenから等距離にある位置に生じた両腕上の切断と誤修復による逆位は、通常法による観察では識別はできません。これは両腕の末端部から等距離のところに生じた切断と再結合でも同じように発見できません。
具体的な例としてまず 図13 を見ることにしましょう。
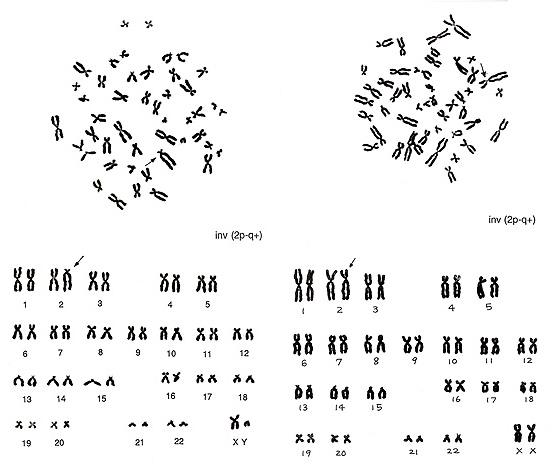
図13.
図13に2番染色体の逆位(矢印)を示します。 左の例は逆位染色体が特異的な形態となっているので、識別は容易です。他の染色体がすべて正常ですから、2番の逆位であることは疑いもありません。右の例は、2番の長腕がやや長く、短腕がやや短いこと、しかも正常2番染色体と異常染色体との間には長さの違いはありません。変化しているのはCIです。
図14は 左がB群染色体、右がC群染色体の逆位です。

図14.
左のB群の例はCIの違いだけが根拠で、短腕のCIが 30%前後です。この程度がおそらく識別しうる限界のように思います。右の図はC群の逆位ですが、異常染色体がD群かG群の「化け物」のような形をしています。長さがC群の範囲にあることと、D群とG群の数がそれぞれ6本と4本であることが確認されれば、C群の逆位であることの判定は容易です。

図15.
図15左は G群が5本(その内の1本はY染色体)、C群が16本(男性だからC群が1本多い)、そしてD群から1本が失われています。C群の過剰の1本は小さい方に属し、長さはD群と変わらないようです。とすれば、 D群の1本が逆位を生じた結果、cenの位置が中央寄りの小型C群染色体へと変化した、と考えるのが妥当のようです。このメタフェースに含まれる染色体の1本が他の染色体と離れた位置にあるのは少々問題です。完全なカリオタイプ分析をしない場合には、染色体数の変異の原因となりそうなメタフェースといえます。
図15. 右は 複雑な異常を持つ例です。まずB群の1本に生じた逆位ですが、その形態は3番染色体に実によく似ています。(下図において矢印をつけた5番染色体)
これを逆位と判定した訳は、逆位染色体は3番よりも長腕、短腕共にやや短いからです。さて、このメタフェースには別の異常も存在しています。即ち、1番によく似た染色体(下図において3番の矢印をつけたもの; hが見られない)と16番によく似た染色体(11番の矢印をつけたもの; 正常の16番よりも短腕が長い)がそれぞれ観察されます。そこでB群染色体の逆位はないものと考えて(即ち5番の矢印をつけたものを正常の3番染色体とみなして)、これら2本の異常染色体のみの組み合わせにより、異常は t(Bq-; Cp+)のみとすることも可能です。
しかし、そうしなかったのはB群の逆位と考えた異常染色体(5番の矢印をつけたもの)と正常の3番染色体との間には長さの違いがあるために、正常のペアとするには問題があると考えたからです。そして、逆位と転座の両方を持つと解釈しました。
第10章.安定型構造異常-各論3.相互転座
転座型異常は原爆被爆者のリンパ球に見られる安定型異常の中でもっとも頻度の高い構造異常で、安定型異常の 約70%を占めています。放射線はランダムにDNAに切断を生じるので、転座の誘発率は染色体の長さにほぼ比例します。
通常法による転座型異常の識別上の難しさは数多くありますが、中でも、2本の染色体の間で交換される染色体片の長さが同じならば、転座として識別することはできません。さらに、C群のように同じような長さで、しかも似たような形態を持つC群染色体の間で生じる転座は実際に起こりうる転座の頻度よりもはるかに低い割合でしか識別されません。残念ながら、これが通常法による異常識別の限界です。
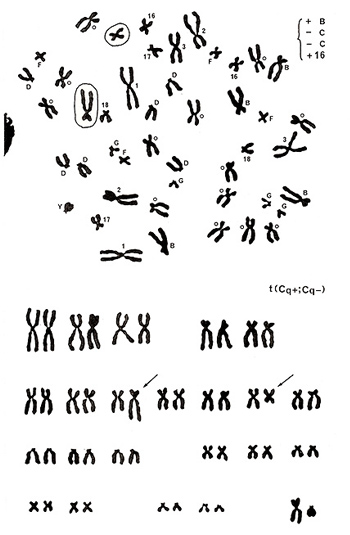
図16.
例を見ることにしましょう。図16. 上ですが、A群、D群、F群、G群およびYには異常がありません。B群は5本、16番が3本とそれぞれ1本ずつ増えている代わりに、C群(男性ですから15本が正常)が2本足りません。
分析の結果(図16. 下)は、C群の1本の長腕が長くなり、B群と非常によく似た形になっていますし、他の1本の長腕が短くなり、長さも形も16番とそっくりです。つまり、C群の2本の染色体の長腕同志の交換(転座)により、一方はB状となり、他方は16番状を示している訳です。かくて、異常のタイプはt(Cq+; Cq-)と決定されました。
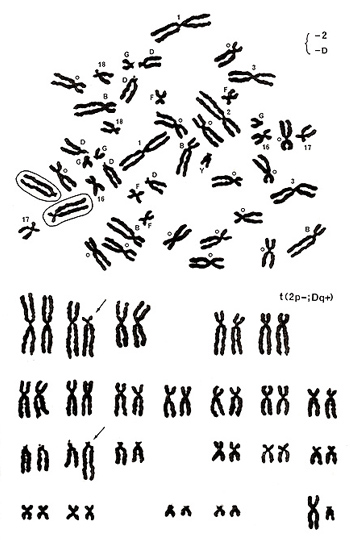
図17.
次に図17. について説明します。メタフェースを見るとD群の1本の長腕が長く、2番の1本が見当たらない代わりに、17番の長腕が著しく伸びたような奇妙な染色体があります。解析結果は、図17下に示したように2番短腕とD群の1本の長腕との転座t(2p-; Dq+)であることがわかります。
図16、17から分かることは、逆位や欠失ではただ1本の染色体の異常に由来しますが、転座の場合は最低2本以上の染色体に生じた交換型の異常ですから、いくつかの染色体群の数に変動が生じています。ここに「欠失・逆位」型異常と「転座」型異常を見分けるコツがあります。
ここでは長い染色体に観察される転座を示しています。異常な染色体はメタフェース、カリオタイプ共に矢印で示してあります。図18 から 図20 までの6例のメタフェースはいずれも1番染色体が関与している異常です。
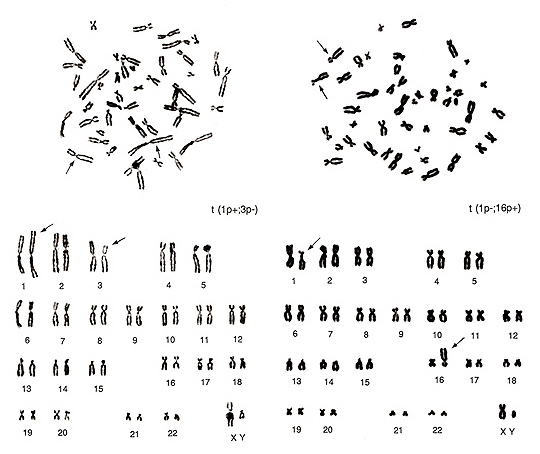
図18.
図18. 左のメタフェースは重なりが多く、また染色体もやや細いので、初心者には異常の判定が難しいかもしれません。
異常はA群に見られます。3番が1本不足し、その代わりに大型のC群染色体が1本余分に存在します。また、1番の一方が長腕、短腕共に長いことが分かります。この染色体をよく見ると、異常の1番の“h”が異様に長い、つまり異形染色体であることがわかります。
これは正常変異の範囲なので、実は短腕が異常に長いのです。余分のC群染色体は3番に属すると考えれば、異常の型は1番短腕と3番短腕の間の転座 t(1p+; 3p-) となります。
図18右では、1番の短腕が短く、かつ長腕のcen付近にかなり大きなhが認められます。E群(16番)の1本は失われ、その代わりにB群が1本余分にあります。転座は当然1番短腕と16番短腕との間に起こったものと判断されます。因みに、この図の2つのメタフェースは同一人物からのものですから、1番のhに関する正常変異が両方の細胞に観察されるのは当然のことです。
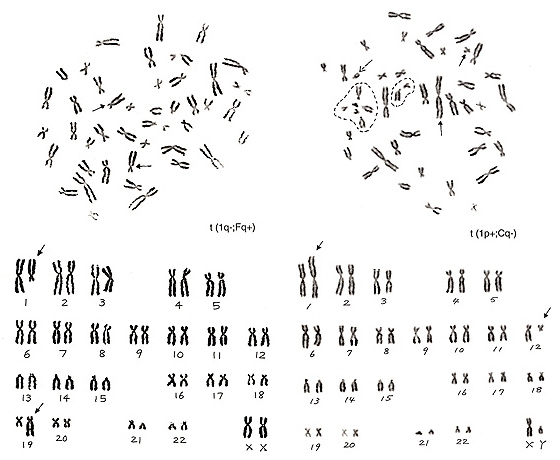
図19.
図19. 左は1番とF群からそれぞれ1本染色体が失われ、その代わりに2本の過剰なC群様の染色体が存在します。
したがって、転座t(1q-; Fq+)となります。図19. 右は、1番の短腕が極端に長く、G群染色体が6本(ただし1本はY)となり、C群が1本不足しています。これらを総合すると、G群様の1本は実はC群に由来するもので、転座のために長腕の大部分が1番の短腕に移ったと考えるのが妥当です。
ではG群の内のどれがC群由来の染色体かを推理します。Y(破線の矢印)を除く5本のG群染色体のうち4本は破線で囲ったサークルの中にあります。これらはD群染色体と共に付随体連合(satellite association)を作っているので、本物のG群染色体です。従って残った1本(矢印)がC群染色体に関わる異常染色体と推定されます。
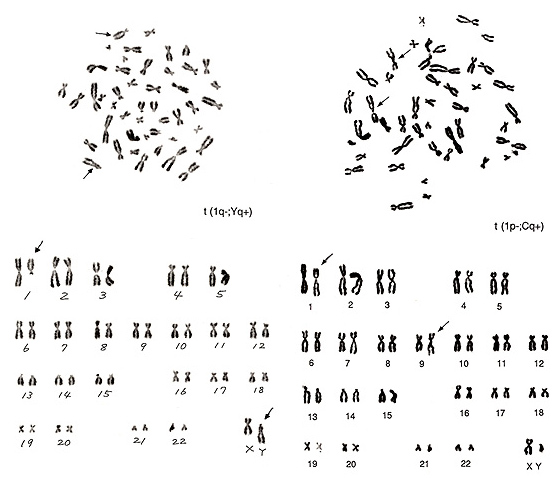
図20.
図20. も1番が関与する転座です。
左は、1番が1本不足し、まるでD群の「お化け」のような染色体が1本あります。通常Y染色体の長腕下部はhが多く、その部分では2本の染色分体はG群染色体よりもくっついているという特徴があります。このお化け染色体の上半分にはYの長腕の特徴がよく現れていますから、1番長腕の多くがYの長腕に転座した、t(1q-; Yq+)という異常が形成されたものと判断されます。
図20. 右の異常にはいろいろな特徴が見られます。まず、1番の短腕が短くなっていること、および、この異常染色体のhはかなり大きいことが分かります。転座のパートナーはC群の1本で、これもhの大きい(恐らく)9番染色体の長腕が長くなっています。したがって、異常はt(1p-; Cq+)となります。
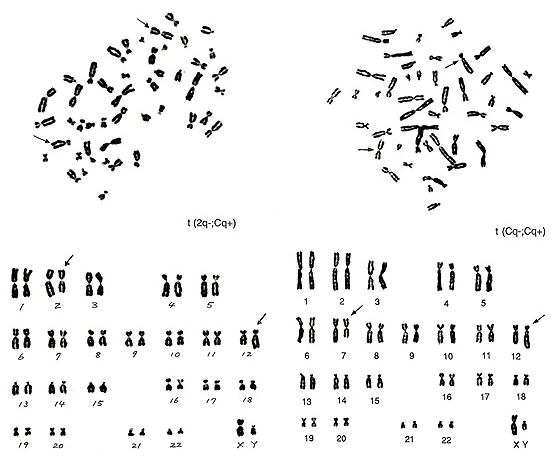
図21.
図21. 左の異常は、A群3番が3本あり、B群染色体が1本余分にあることに基づくものです。
これら2本の正常様の異常染色体がメタフェースの観察だけで識別されたなら、あなたはもう素人の域を脱したと思われます。それから後の異常の判定は簡単で、図にもある通り、t(2q-; Cq+)です。図21. 右は判定がもっとも難しいC群染色体の間の転座です。矢印で示す異常染色体は、一方は小型でcenが染色体の中央に位置し、他方はB群によく似ているが、短腕が小型のもの、の2本からなります。
いろいろな異常を想定できますが、t(Cq-; Cp+)というのが妥当なところです。
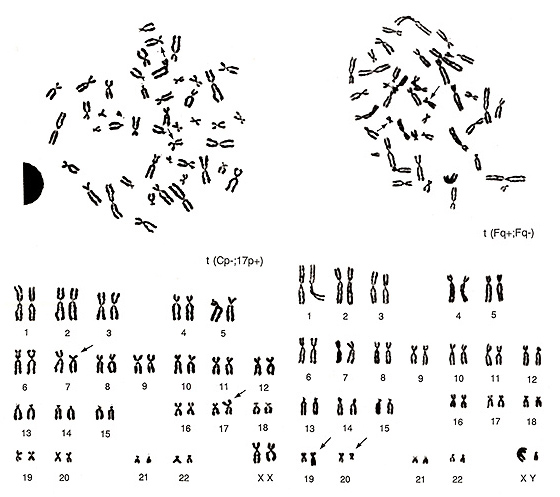
図22.
図22. には比較的小型の染色体が関与した転座例を示します。
左図の異常はメタフェースのレベルではなかなか難しいものです。よく観察すると、E群の1本(多分17番)が欠けており、C群が1本余計にあります。小型のC群様の染色体はcenがほぼ中央部にありますから、異常には間違いありません。中型のC群染色体の1本はCIのバランス(短腕が短く、長腕が長い)が悪く、“C群らしくない”ことに注目すべきです。カリオタイプの結果はt(Cp-; 17p+)とするのがもっとも抵抗が少ないと思います。
図22. 右は、F群が2本少なく、16番とも17番ともつかない染色体が1本と、G群よりも小型の染色体が観察されます。カリオタイプからも分かるように、2本のF群染色体の転座t(Fq+; Fq-)です。
最後の例を見ても分かるように、F群やG群のように小さな染色体の異常では、染色体の増減がほんのわずかであってもかなり的確に把握することができます。従って、通常法で安定型異常を識別することは、大型の染色体が関与する異常を見逃す結果となるのは避けられません。
第11章.誤った判定の例
最後に誤りの例を紹介します。図23. を見て下さい。左側のメタフェースは、実は 図6 左に示すように正常染色体の代表例なのです。図23. の右上にその正常パターンを示します。
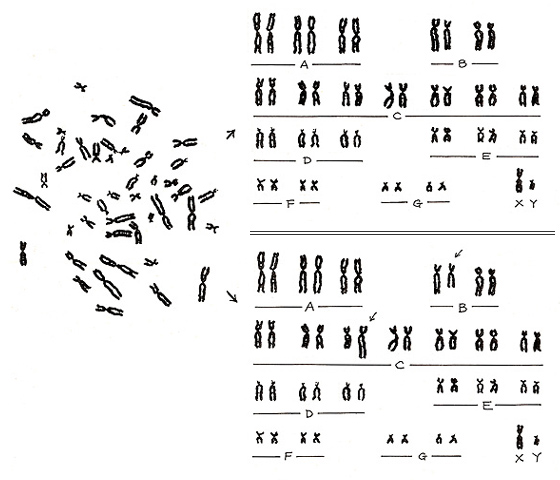
図23.
しかし、どこかに誤解を生じると、図23右下にあるような異常パターンになりかねない、ということです。正常パターンでも異なる染色体群の中から1本ずつ取り出して入れ替えると、異常パターンにしてしまうことが可能なのです。経験を積み重ねることによって、このような誤りを少なくすることは可能です。しかし、どこに落とし穴が口をあけて待っているか分かりません。疑いをもったメタフェースは必ず写真撮影をし、後日の証拠に残しておくという慎重な態度が望まれます。
第12章.終わりに
明治維新に活躍し、「天は人の上に人を造らず、人の下に人を造らずといえり」の名文句で知られ、そして慶応義塾大学の創始者でもある福澤諭吉が書いた「通俗医術論」という著書の中に次の一節があります。
「蓋(けだ)し固有(もちまえ)は常なり、疾病は変なり。固有を知らざれば、則(すなわ)ち病変を論ず可からざるなり」 (土屋雅春著、“医者の見た福澤諭吉”中公新書、115頁、1996年)。この内容はごく当たり前のことをいっているに過ぎません。この一文にある“固有(もちまえ)”を正常染色体に、また、“疾病”や“病変”を異常染色体に読み替えてみると、「正常染色体を知らないで、なんで異常染色体を論ずることができるか。」となります。私には福澤諭吉が「正常を知らずして異常を語るなかれ」と、物事を学ぶ上での基本的な心構えについて諭しているように思えるのです。
これは何も福澤諭吉一人に限りません。「きちんとした仕事をするのに本当に必要なのは、ほんのわずかな正しい基本動作をきちんと行うことで充分足りる。」 これはアメリカのゴルフの名人・達人と呼ばれた人が、自分の一生を振り返って述懐した言葉です。
基本とは何も特別なものではありません。簡単で当たり前のことなのです。基本を学ぶこと自体は別に難しいわけではありません。しかし、それを実行するのは意外に難しいのかもしれません。
この手引書で繰り返し述べてきたように、染色体異常は多種多様ですが、正常パターンは一種類しかありません。正常を熟知していなければ、異常の識別はできないのです。正常パターンを身につけるのに多少のコツはあっても、残念ながら近道はありません。何事も基本が大事なのですが、基本をマスターするためには忍耐が必要です。しかしそれが可能であると分かっているだけ希望はあります。ですからこの基本をマスターすることの重要さだけは是非記憶し、心に刻み込んで戴きたいのです。
最後になりました。人類染色体数が46であることを最初に発表したのはJ. H. Tjio(チヨー)博士とA. Levan(レヴァン)博士で、1956年のことです。今年ですでに44年も経ちました。その頃私は大学院修士課程の1年に入学したばかりでした。それから1、2年経ってからこの論文を初めて読み、それまでのヒト染色体数についての47説や48説がいずれも間違いであったことを知って驚きました。私は1958年から1993年迄の35年間、ヒトの染色体に関わる研究に携わってきました。1995年で研究生活を事実上終えましたが、それでも今日迄を通算すると40年になります。
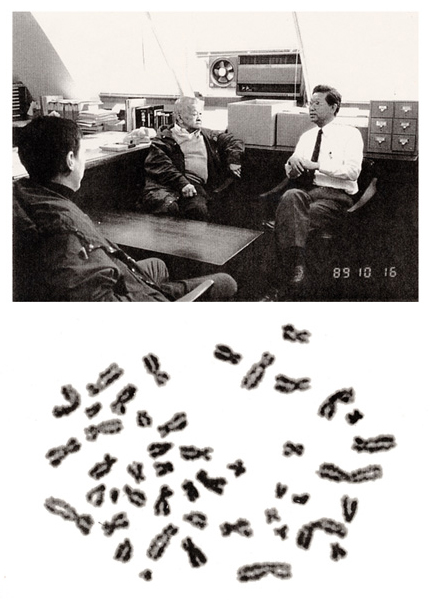
図24.
1989年10月、上記のチヨー博士が家族と共に初めて放影研を訪問されました。チヨー先生は永年米国のNIHで研究生活をされておりました。うわさによればなかなか気難しい方だと伺っておりましたが、会ってお話をするうちに、気さくで子煩悩な1人の父親であることが次第に分かってきました。最初のころは私の方が緊張しておりましたが、私の部屋での話しも和やかな雰囲気の中で一段と弾んでゆきました。帰国後、チヨー先生から感謝の手紙を戴きましたが、その中に、1956年に発表した論文に引用されているメタフェースの染色体を拡大した顕微鏡写真が同封されていました。
この時に撮影したチヨー先生の写真をここに示します。(図24. 上、写真中央の人物)
また、図24. 下の写真はスエーデンの遺伝学専門誌 “Hereditas” に掲載されたヒトのメタフェースで、染色体数が46本であることを世界で最初に示した時の写真を拡大プリントして送られてきたものです。これら2枚の写真を本文の最後に飾りたいと思います。